
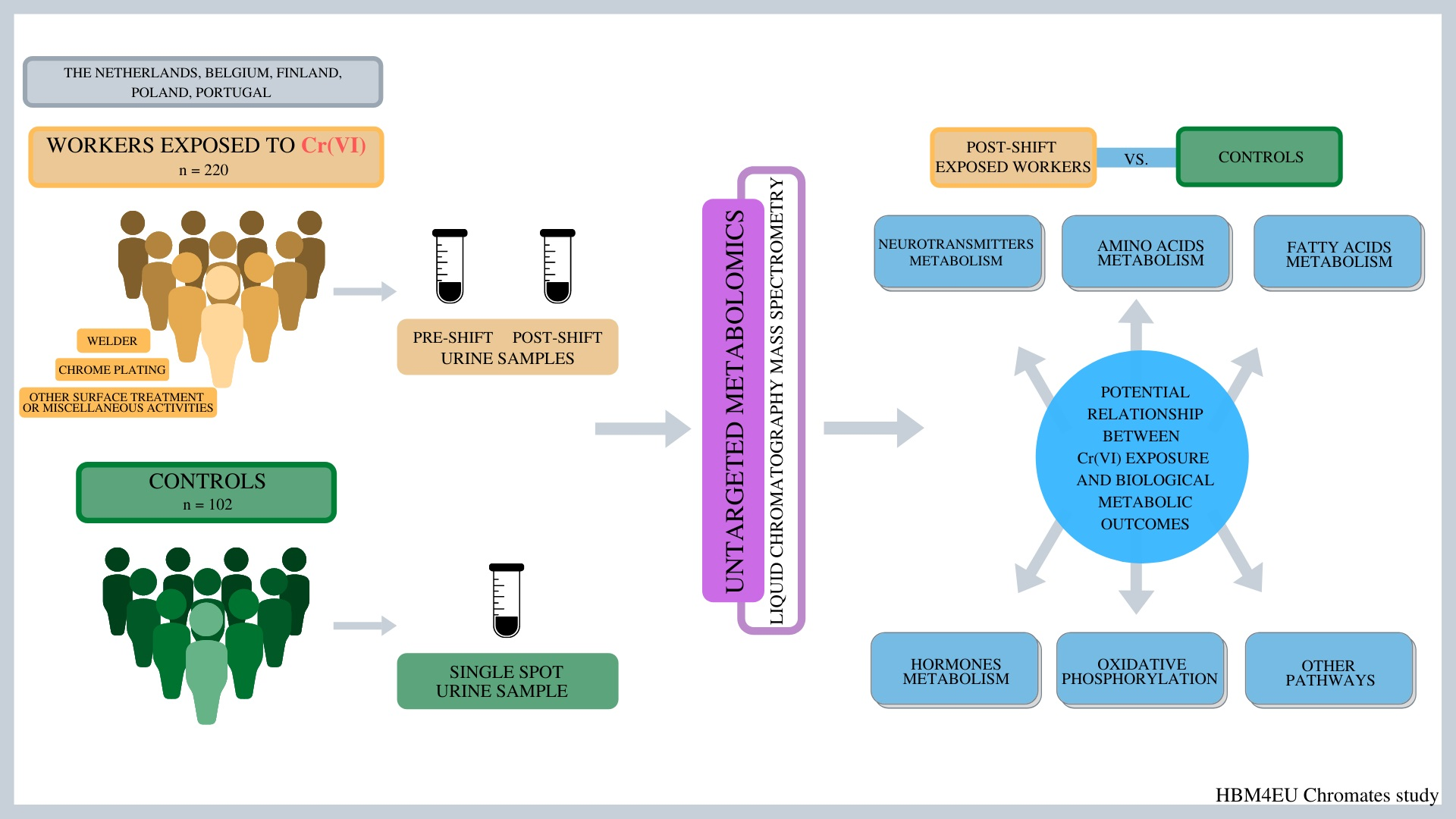
HBM4EU Chromates Study: Urinary Metabolomics Study of Workers Exposed to Hexavalent Chromium

 ,
,  ,
,  ,
,  ,
,  ,
,  , , , ,
, , , ,  , ,
, ,
Abstract

1. Introduction
2. Results
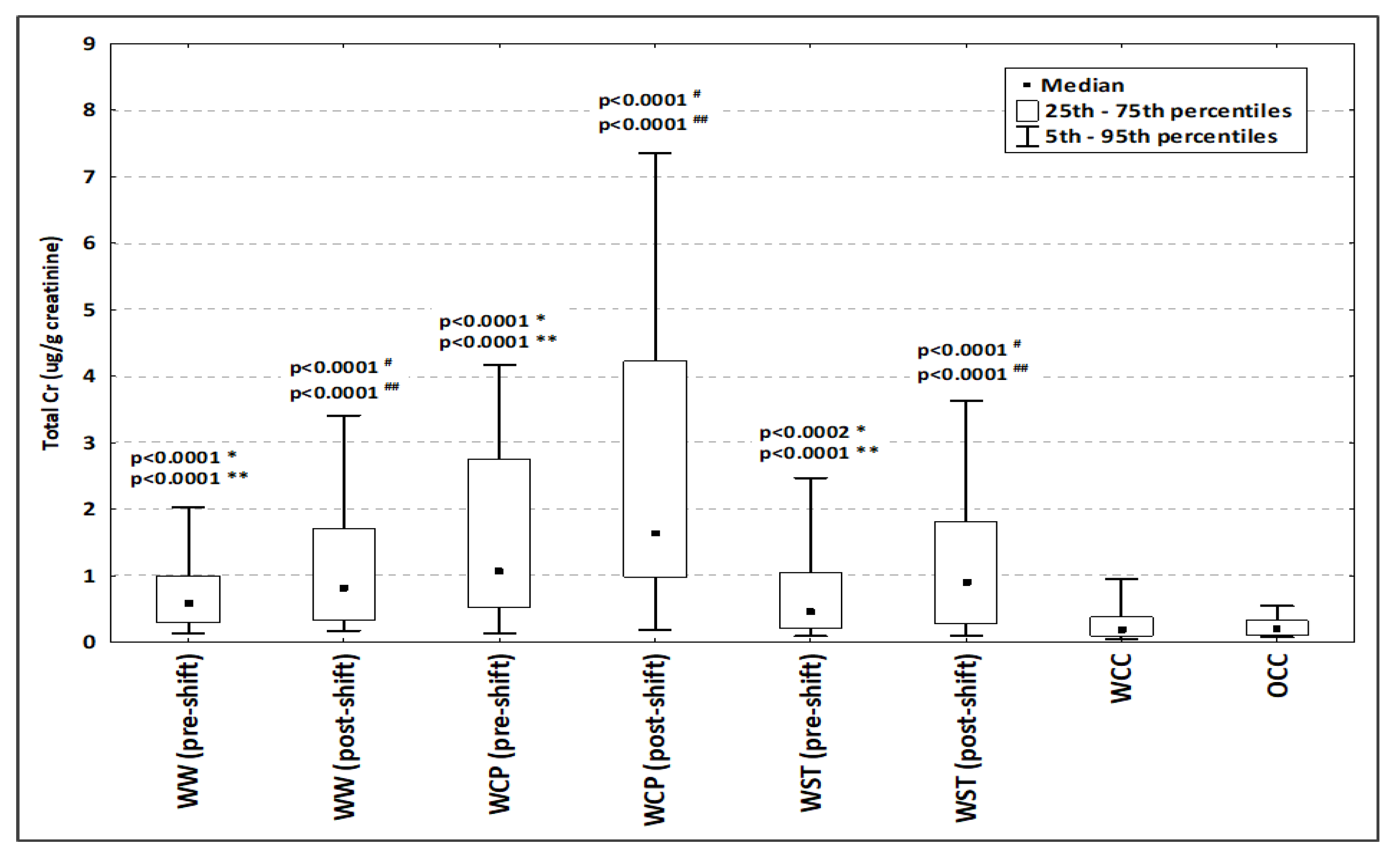
2.1. General Characteristics of the Study Population
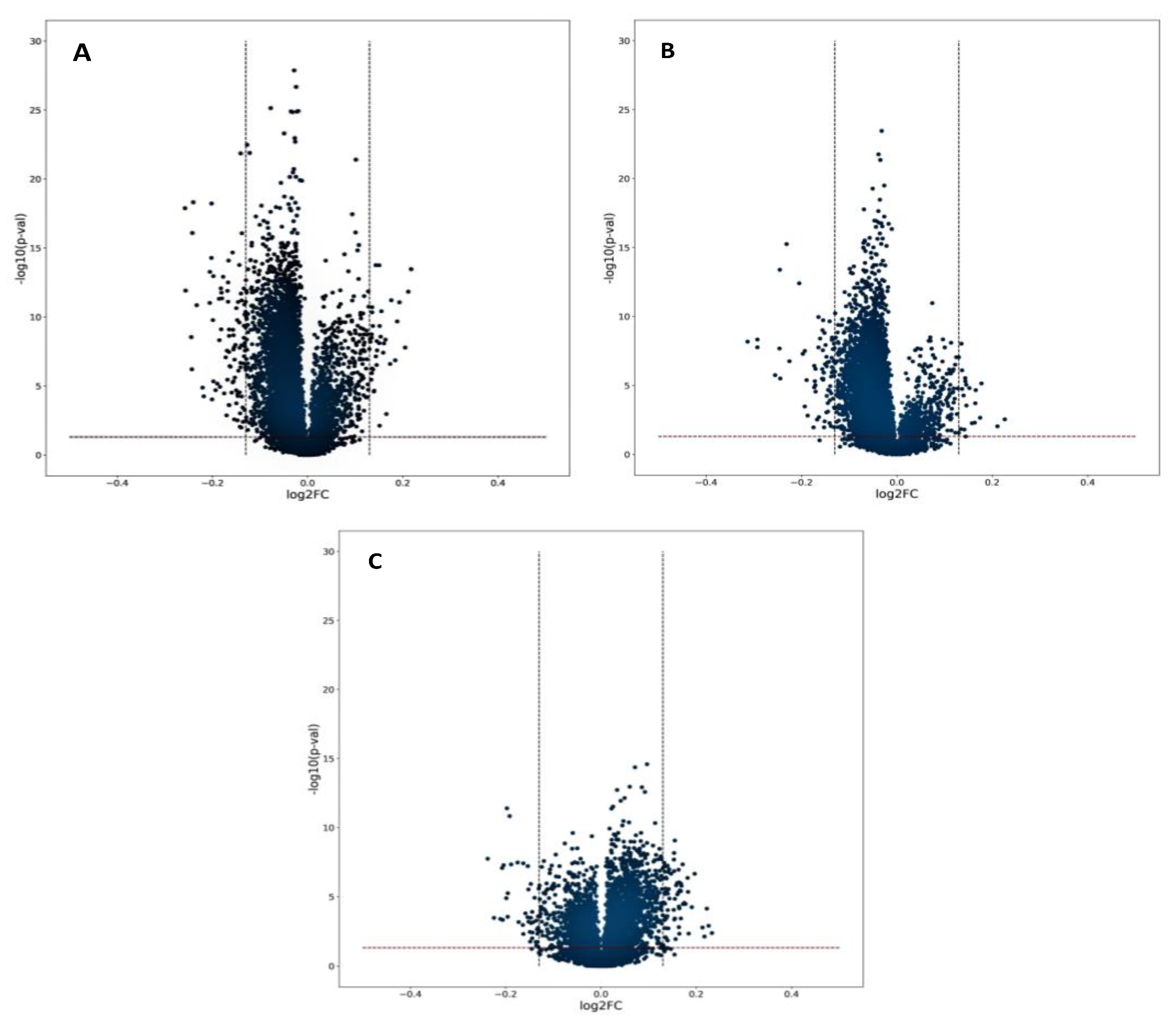
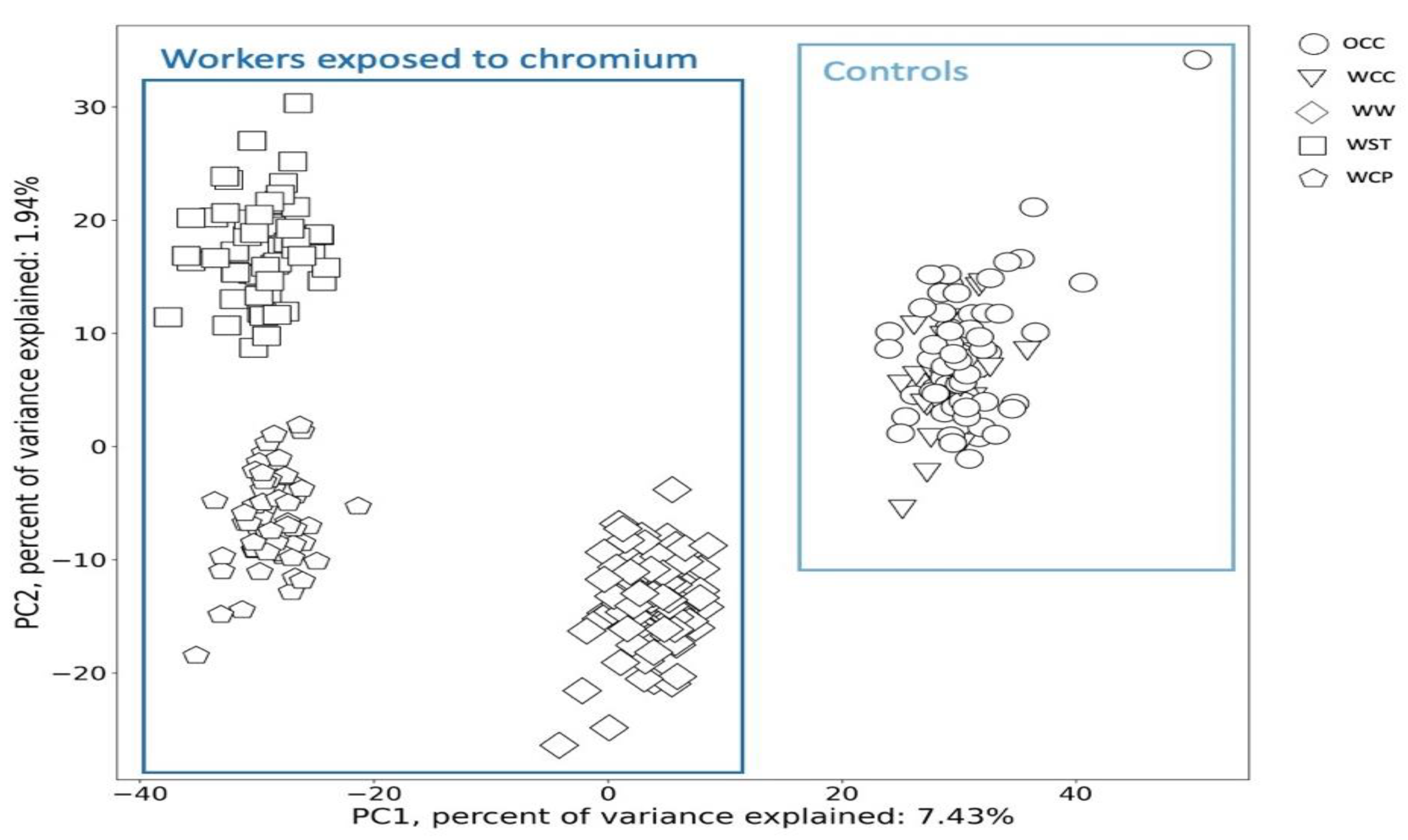
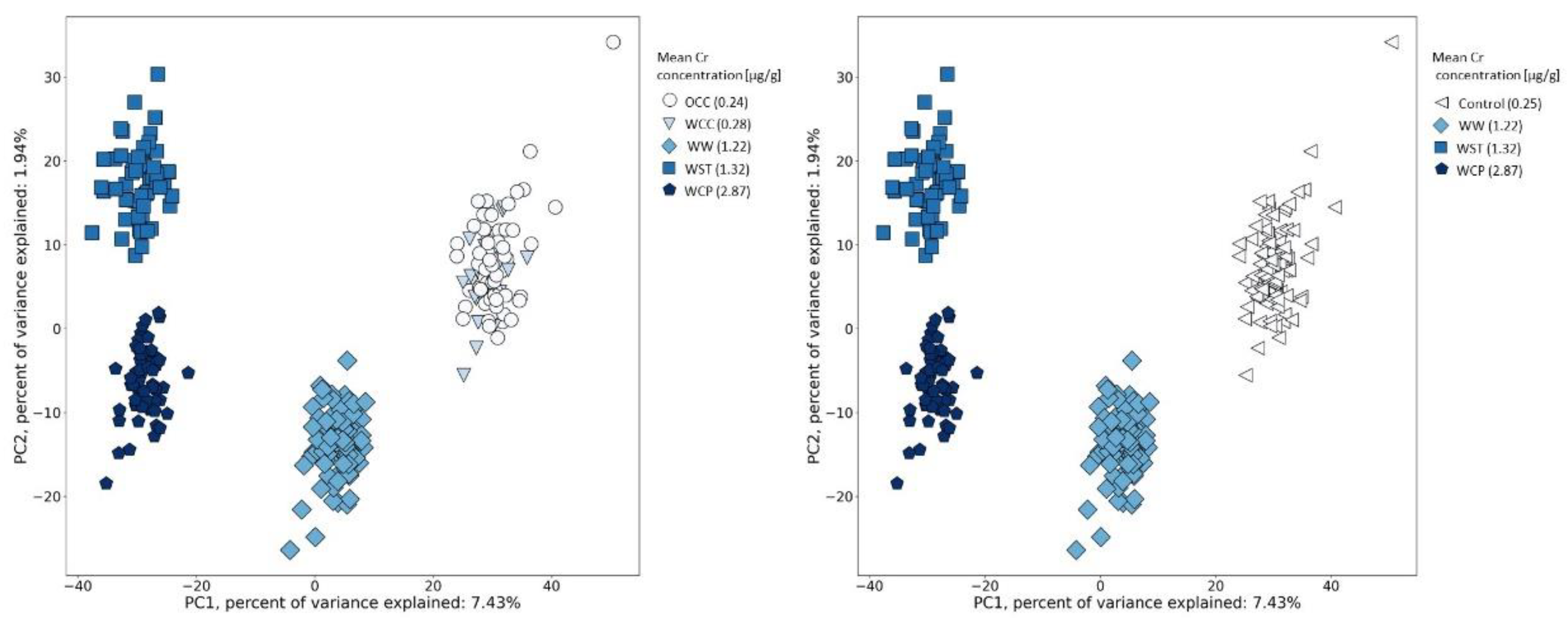
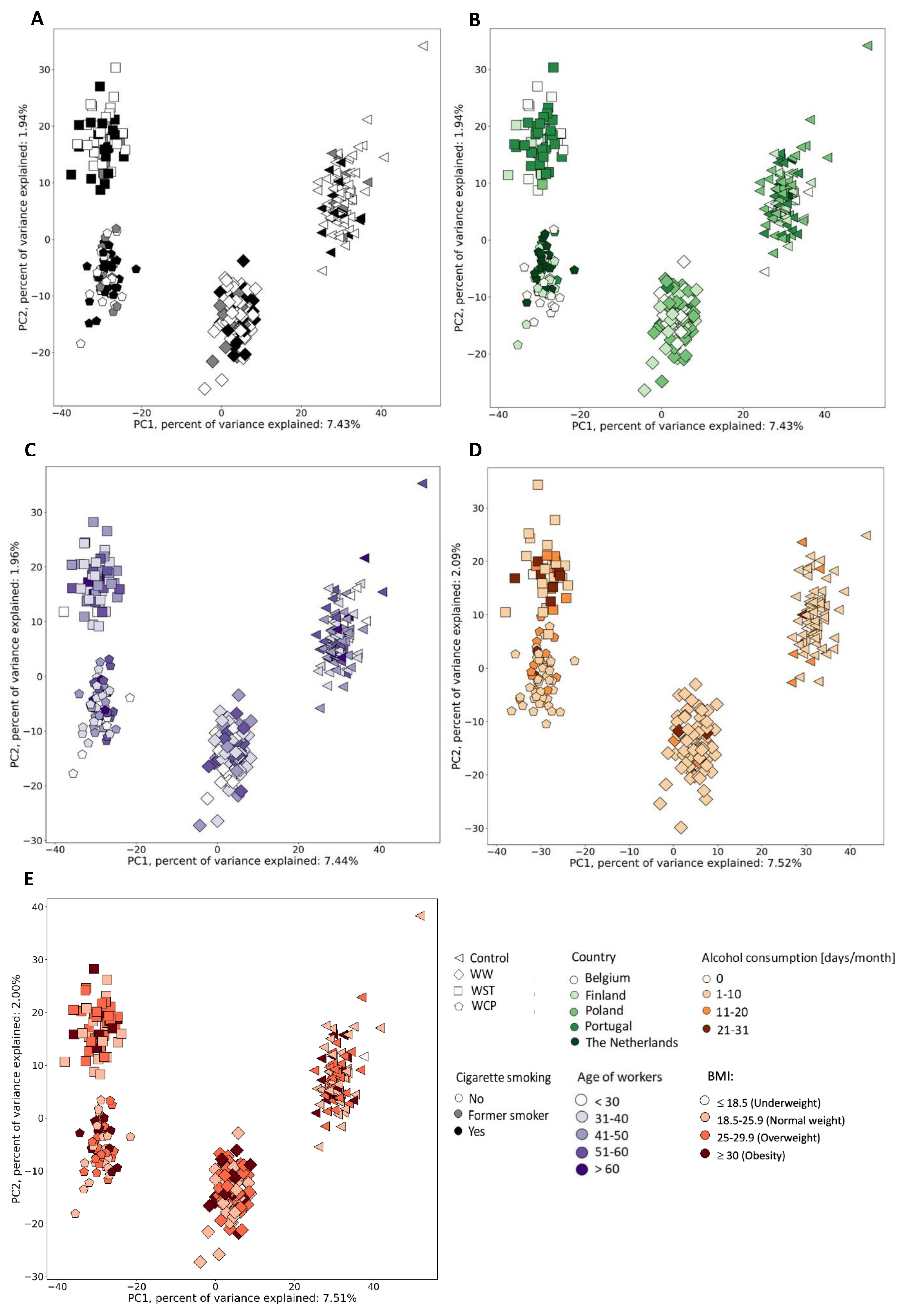
2.2. Urine Metabolic Changes following Cr(VI) Exposure
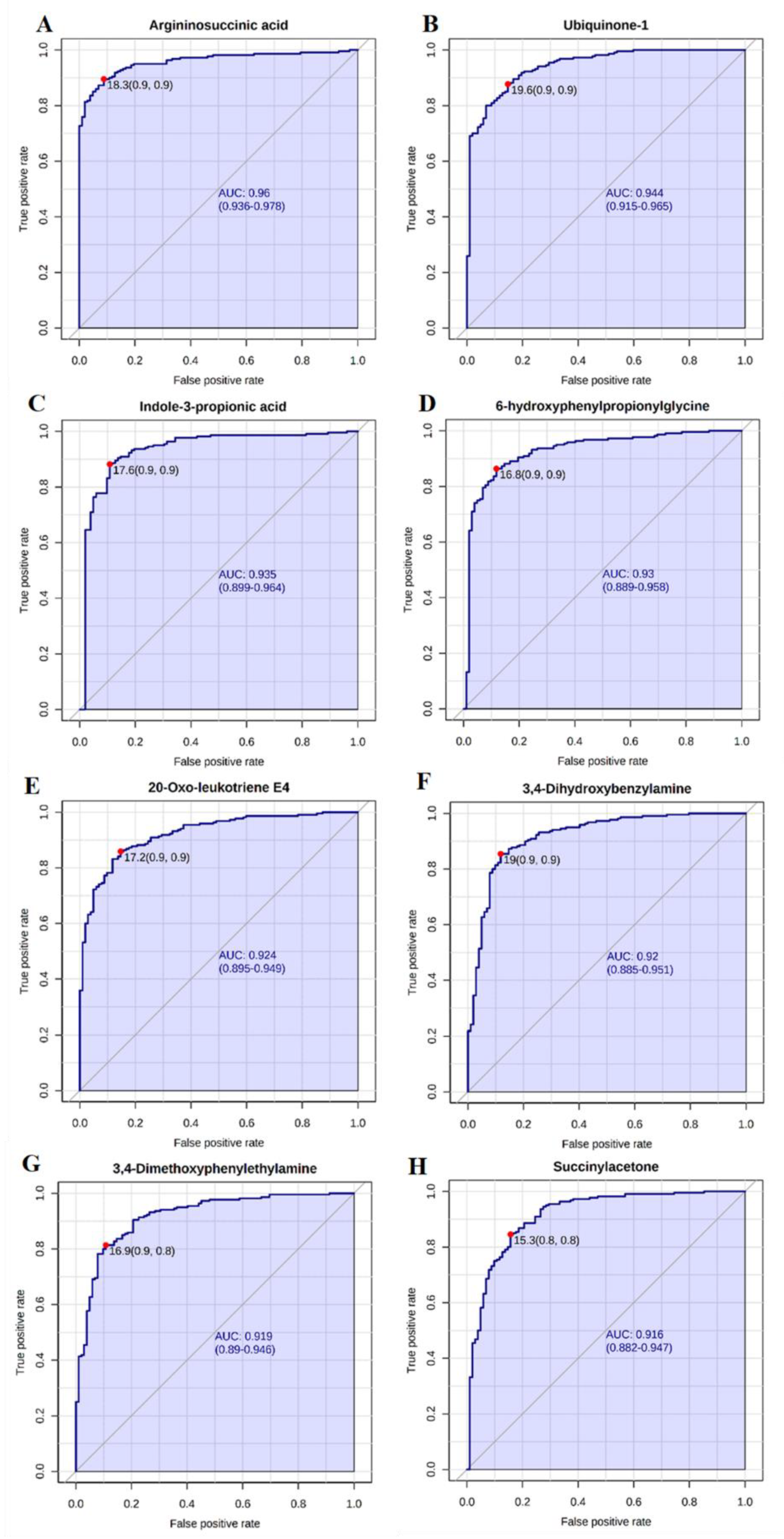
2.3. Candidate Biomarkers of Occupational Exposure to Cr(VI)
3. Discussion
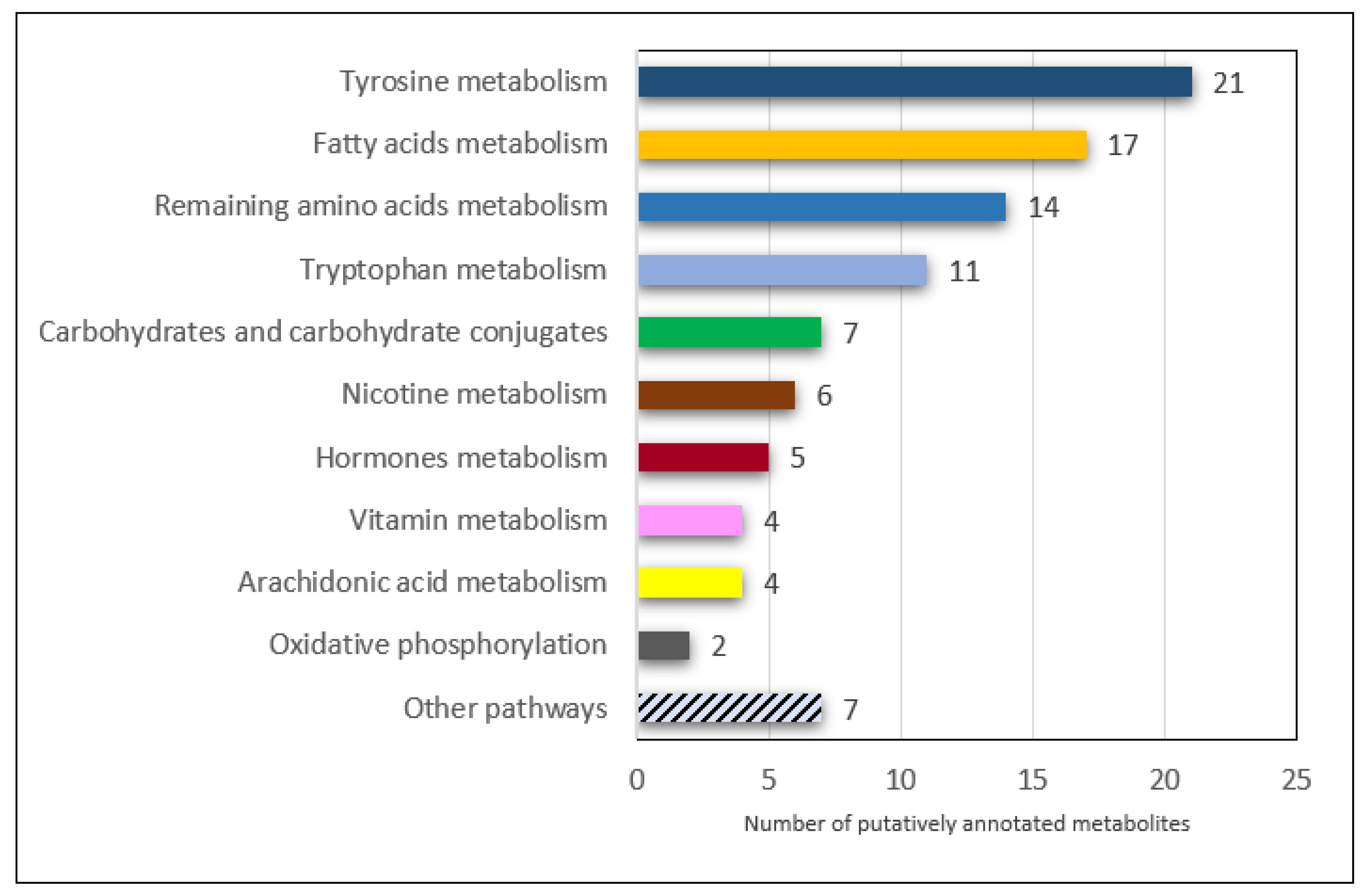
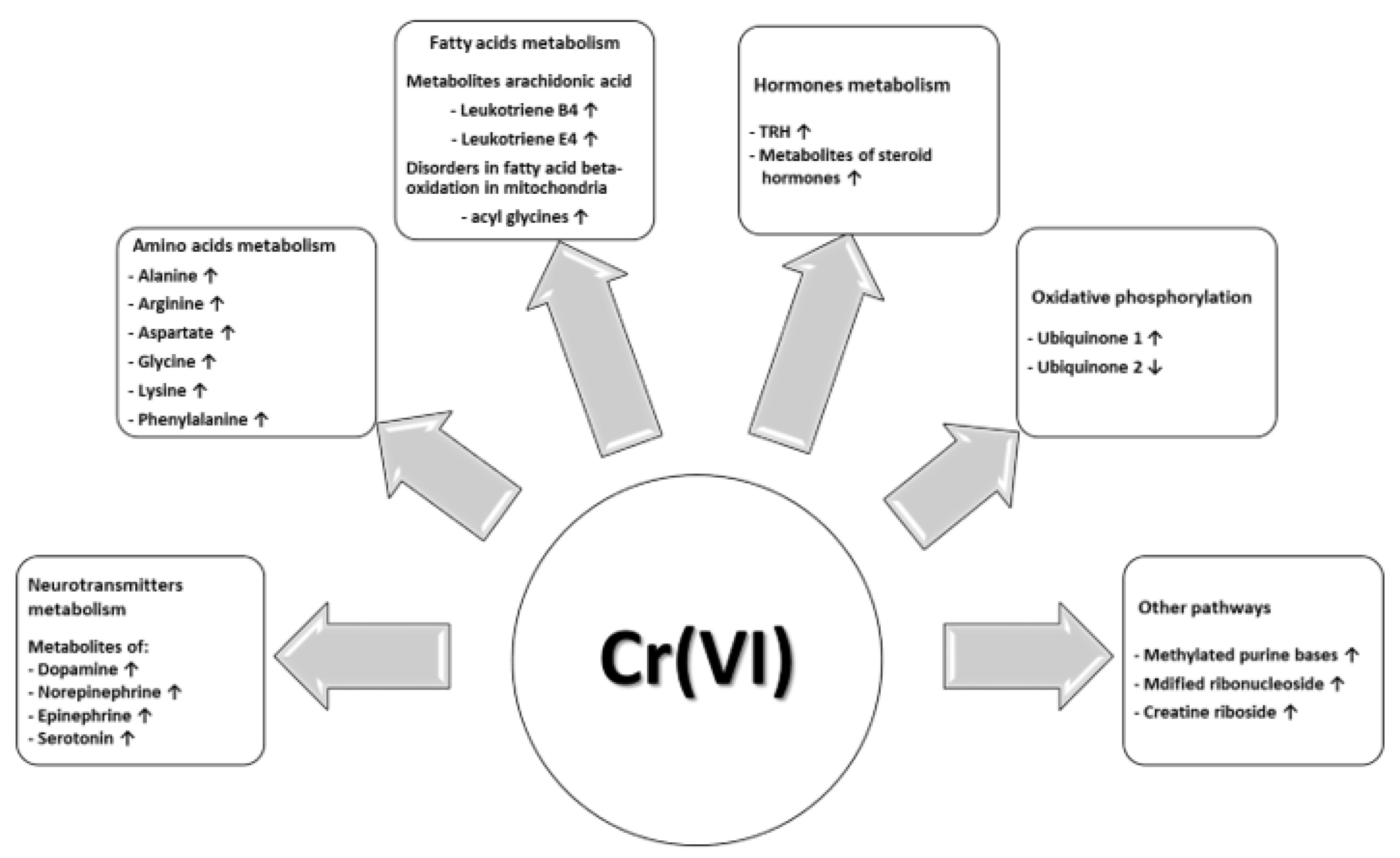
3.1. Tyrosine, Tryptophan and Other Amino Acid Metabolism
3.2. Arachidonic Acid and Other Fatty Acids Metabolism
3.3. Hormones Metabolism
3.4. Oxidative Phosphorylation
3.5. Nicotine, Vitamins and Other Pathways
3.6. Candidate Biomarkers of Occupational Exposure to Cr(VI)
3.7. Strengths and Limitations of the Study
4. Materials and Methods
4.1. Study Population
4.2. Urine Collection and Cr Analysis
4.3. Sample Preparations for Untargeted Metabolomics
4.4. Metabolomics Analysis
4.5. Metabolomic Data Preoprocessing and Normalization
4.6. Statistical and Chemometric Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- National Institute for Occupational Safety and Health (NIOSH). Occupational Exposure to Hexavalent Chromium; U.S. Department of Health and Human Services (DHHS), NIOSH: Washington, DC, USA, 2013; pp. 1–148. [Google Scholar]
- Aitio, A.; Jarvisalo, J.; Kiilunen, M.; Kalliomaki, P.L.; Kalliomaki, K. Chromium. In Biological Monitoring of Toxic Metals; Clarkson, T.W., Friberg, L., Nordberg, G.F., Sager, P.R., Eds.; Springer US: Boston, MA, USA, 1988; pp. 369–382. [Google Scholar] [CrossRef]
- Alvarez, C.C.; Bravo Gómez, M.E.; Hernández Zavala, A. Hexavalent Chromium: Regulation and Health Effects. J. Trace Elem. Med. Biol. 2021, 65, 126729. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Evaluation of Carcinogenic Risks to Humans. Chromium, Nickel and Welding; IRAC: Lyon, France, 1990; pp. 1–687. [Google Scholar]
- Borthiry, G.R.; Antholine, W.E.; Kalyanaraman, B.; Myers, J.M.; Myers, C.R. Reduction of Hexavalent Chromium by Human Cytochrome B5: Generation of Hydroxyl Radical and Superoxide. Free Radic. Biol. Med. 2007, 42, 738–755. [Google Scholar] [CrossRef]
- Hu, G.; Zheng, P.; Feng, H.; Jia, G. Imbalance of Oxidative and Reductive Species Involved in Chromium(VI)-Induced Toxic Effects. ROS 2017, 3, 1–11. [Google Scholar] [CrossRef]
- O’Brien, T. Complexities of Chromium Carcinogenesis: Role of Cellular Response, Repair and Recovery Mechanisms. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2003, 533, 3–36. [Google Scholar] [CrossRef]
- Chen, Q.Y.; DesMarais, T.; Costa, M. Metals and Mechanisms of Carcinogenesis. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 537–554. [Google Scholar] [CrossRef]
- Lankford, E.; Thomas, C.S.; Marchi, S.; Brown, S.; Woski, S.A.; Vincent, J.B. Examining the Potential Formation of Ternary Chromium-Histidine-DNA Complexes and Implications for Their Carcinogenicity. Biol. Trace Elem. Res. 2022, 200, 1473–1481. [Google Scholar] [CrossRef]
- Agency for Toxic Substances and Disease Registry (ATSDR). Toxicological Profile for Chromium; Department of Health and Human Services, Public Health Service: Atlanta, GA, USA, 2000; pp. 1–592. [Google Scholar]
- International Agency for Research on Cancer (IARC). Arsenic, Metals, Fibres, and Dust, Volume 100 C, a Review of Human Carcinogens; IRAC: Lyon, France, 2012; pp. 1–527. [Google Scholar]
- Lindberg, E.; Hedenstierna, G. Chrome Plating: Symptoms, Findings in the Upper Airways, and Effects on Lung Function. Arch. Environ. Health 1983, 38, 367–374. [Google Scholar] [CrossRef]
- Dayan, A.D.; Paine, A.J. Mechanisms of Chromium Toxicity, Carcinogenicity and Allergenicity: Review of the Literature from 1985 to 2000. Hum. Exp. Toxicol. 2001, 20, 439–451. [Google Scholar] [CrossRef]
- Li, H.; Chen, Q.; Li, S.; Yao, W.; Li, L.; Shi, X.; Wang, L.; Castranova, V.; Vallyathan, V.; Ernst, E.; et al. Effect of Cr(VI) Exposure on Sperm Quality: Human and Animal Studies. Ann. Occup. Hyg. 2001, 45, 505–511. [Google Scholar] [CrossRef]
- Bruynzeel, D.P.; Hennipman, G.; van Ketel, W.G. Irritant Contact Dermatitis and Chrome-Passivated Metal. Contact Dermat. 1988, 19, 175–179. [Google Scholar] [CrossRef]
- Sharma, B.K.; Singhal, P.C.; Chugh, K.S. Intravascular Haemolysis and Acute Renal Failure Following Potassium Dichromate Poisoning. Postgrad. Med. J. 1978, 54, 414–415. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, E.; Vesterberg, O. Urinary Excretion of Proteins in Chromeplaters, Exchromeplaters and Referents. Scand. J. Work Environ. Health 1983, 9, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Ganzleben, C.; Antignac, J.P.; Barouki, R.; Castaño, A.; Fiddicke, U.; Klánová, J.; Lebret, E.; Olea, N.; Sarigiannis, D.; Schoeters, G.R.; et al. Human Biomonitoring as a Tool to Support Chemicals Regulation in the European Union. Int. J. Hyg. Environ. Health 2017, 220, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Santonen, T.; Porras, S.P.; Bocca, B.; Bousoumah, R.; Duca, R.C.; Galea, K.S.; Godderis, L.; Göen, T.; Hardy, E.; Iavicoli, I.; et al. HBM4EU Chromates Study—Overall Results and Recommendations for the Biomonitoring of Occupational Exposure to Hexavalent Chromium. Environ. Res. 2022, 204, 111984. [Google Scholar] [CrossRef]
- Ventura, C.; Gomes, B.C.; Oberemm, A.; Louro, H.; Huuskonen, P.; Mustieles, V.; Fernández, M.F.; Ndaw, S.; Mengelers, M.; Luijten, M.; et al. Biomarkers of Effect as Determined in Human Biomonitoring Studies on Hexavalent Chromium and Cadmium in the Period 2008–2020. Environ. Res. 2021, 197, 110998. [Google Scholar] [CrossRef]
- Beaulieu, J.M.; Espinoza, S.; Gainetdinov, R.R. Dopamine Receptors—IUPHAR Review 13: Dopamine Receptors. Br. J. Pharmacol. 2015, 172, 1–23. [Google Scholar] [CrossRef]
- Li, M.; Zhou, L.; Sun, X.; Yang, Y.; Zhang, C.; Wang, T.; Fu, F. Dopamine, a Co-Regulatory Component, Bridges the Central Nervous System and the Immune System. Biomed. Pharmacother. 2022, 145, 112458. [Google Scholar] [CrossRef]
- Mezzelani, A.; Landini, M.; Facchiano, F.; Raggi, M.E.; Villa, L.; Molteni, M.; De Santis, B.; Brera, C.; Caroli, A.M.; Milanesi, L.; et al. Environment, Dysbiosis, Immunity and Sex-Specific Susceptibility: A Translational Hypothesis for Regressive Autism Pathogenesis. Nutr. Neurosci. 2015, 18, 145–161. [Google Scholar] [CrossRef]
- Charnay, Y.; Léger, L. Brain Serotonergic Circuitries. Dialogues Clin. Neurosci. 2010, 12, 471–487. [Google Scholar] [CrossRef]
- Marchitti, S.A.; Deitrich, R.A.; Vasiliou, V. Neurotoxicity and Metabolism of the Catecholamine-Derived 3,4-Dihydroxyphenylacetaldehyde and 3,4-Dihydroxyphenylglycolaldehyde: The Role of Aldehyde Dehydrogenase. Pharmacol. Rev. 2007, 59, 125–150. [Google Scholar] [CrossRef]
- Wu, L.; Gong, W.; Shen, S.P.; Wang, Z.H.; Yao, J.X.; Wang, J.; Yu, J.; Gao, R.; Wu, G. Multiple Metal Exposures and Their Correlation with Monoamine Neurotransmitter Metabolism in Chinese Electroplating Workers. Chemosphere 2017, 182, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, L.; Janasik, B.; Nowicka, K.; Wąsowicz, W. A Urinary Metabolomics Study of a Polish Subpopulation Environmentally Exposed to Arsenic. J. Trace Elem. Med. Biol. 2019, 54, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Kiehn, J.T.; Faltraco, F.; Palm, D.; Thome, J.; Oster, H. Circadian Clocks in the Regulation of Neurotransmitter Systems. Pharmacopsychiatry 2019. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.C.; Preto, S. Should an Artificial Window Substitute a Natural One? In Advances in Usability and User Experience; Ahram, T., Falcão, C., Eds.; Advances in Intelligent Systems and Computing; Springer International Publishing: Cham, Switzerland, 2018; Volume 607, pp. 247–258. [Google Scholar] [CrossRef]
- Amoretti, M.; Amsler, C.; Bonomi, G.; Bouchta, A.; Bowe, P.; Carraro, C.; Cesar, C.L.; Charlton, M.; Collier, M.J.T.; Doser, M.; et al. Production and Detection of Cold Antihydrogen Atoms. Nature 2002, 419, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, K.S. Oxidative DNA and Protein Damage in Metal-Induced Toxicity and Carcinogenesis. Free Radic. Biol. Med. 2002, 32, 958–967. [Google Scholar] [CrossRef]
- Stadtman, E.R. Oxidation of Free Amino Acids and Amino Acid Residues in Proteins B by Radiolysis and by Metal-Catalyzed Reactions. Annu. Rev. Biochem. 1993, 62, 797–821. [Google Scholar] [CrossRef] [PubMed]
- Sass, J.O.; Mohr, V.; Olbrich, H.; Engelke, U.; Horvath, J.; Fliegauf, M.; Loges, N.T.; Schweitzer-Krantz, S.; Moebus, R.; Weiler, P.; et al. Mutations in ACY1, the Gene Encoding Aminoacylase 1, Cause a Novel Inborn Error of Metabolism. Am. J. Hum. Genet. 2006, 78, 401–409. [Google Scholar] [CrossRef]
- Tanaka, H.; Sirich, T.L.; Plummer, N.S.; Weaver, D.S.; Meyer, T.W. An Enlarged Profile of Uremic Solutes. PLoS ONE 2015, 10, e0135657. [Google Scholar] [CrossRef]
- Cassiman, D.; Zeevaert, R.; Holme, E.; Kvittingen, E.A.; Jaeken, J.A. Novel Mutation Causing Mild, Atypical Fumarylacetoacetase Deficiency (Tyrosinemia Type I): A Case Report. Orphanet J. Rare Dis. 2009, 4, 28. [Google Scholar] [CrossRef][Green Version]
- Zhao, X.Y.; Wang, J.W.; Yin, Y.; Li, K.; Zhang, M.; Yan, F.P. Effect of Tong Xie Yao Fang on Endogenous Metabolites in Urine of Irritable Bowel Syndrome Model Rats. WJG 2019, 25, 5134–5151. [Google Scholar] [CrossRef]
- Kodama, H.; Zhang, J.; Sugahara, K. Novel Priming Compounds of Cystathionine Metabolites on Superoxide Generation in Human Neutrophils. Biochem. Biophys. Res. Commun. 2000, 269, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Epstein, F.H.; Lewis, R.A.; Austen, K.F.; Soberman, R.J. Leukotrienes and Other Products of the 5-Lipoxygenase Pathway: Biochemistry and Relation to Pathobiology in Human Diseases. N. Engl. J. Med. 1990, 323, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Harizi, H.; Corcuff, J.B.; Gualde, N. Arachidonic-Acid-Derived Eicosanoids: Roles in Biology and Immunopathology. Trends Mol. Med. 2008, 14, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Capra, V.; Bäck, M.; Barbieri, S.S.; Camera, M.; Tremoli, E.; Rovati, G.E. Eicosanoids and Their Drugs in Cardiovascular Diseases: Focus on Atherosclerosis and Stroke. Med. Res. Rev. 2013, 33, 364–438. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; DuBois, R.N. Eicosanoids and Cancer. Nat. Rev. Cancer 2010, 10, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Miyata, J.; Arita, M.; Fukunaga, K.; Kawana, A. Current State and Future Prospect of the Therapeutic Strategy Targeting Cysteinyl Leukotriene Metabolism in Asthma. Respir. Investig. 2019, 57, 534–543. [Google Scholar] [CrossRef]
- Walters, G.I.; Moore, V.C.; Robertson, A.S.; Burge, C.B.S.G.; Vellore, A.D.; Burge, P.S. An Outbreak of Occupational Asthma Due to Chromium and Cobalt. Occup. Med. 2012, 62, 533–540. [Google Scholar] [CrossRef]
- Zhong, X.; de Cássia da Silveira e Sá, R.; Zhong, C. Mitochondrial Biogenesis in Response to Chromium (VI) Toxicity in Human Liver Cells. Int. J. Mol. Sci. 2017, 18, 1877. [Google Scholar] [CrossRef]
- Das, S.; Reddy, R.C.; Chadchan, K.S.; Patil, A.J.; Biradar, M.S.; Das, K.K. Nickel and Oxidative Stress: Cell Signaling Mechanisms and Protective Role of Vitamin C. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 1024–1031. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Carneiro-Freire, N.; Seco-Filgueira, M.; Fernández-Fernández, C.; Mouriño-Bayolo, D. Mitochondrial β-Oxidation of Saturated Fatty Acids in Humans. Mitochondrion 2019, 46, 73–90. [Google Scholar] [CrossRef]
- Xiao, F.; Li, Y.; Luo, L.; Xie, Y.; Zeng, M.; Wang, A.; Chen, H.; Zhong, C. Role of Mitochondrial Electron Transport Chain Dysfunction in Cr(VI)-Induced Cytotoxicity in L-02 Hepatocytes. Cell Physiol. Biochem. 2014, 33, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- García-Niño, W.R.; Zazueta, C.; Tapia, E.; Pedraza-Chaverri, J. Curcumin Attenuates Cr(VI)-Induced Ascites and Changes in the Activity of Aconitase and F(1)F(0) ATPase and the ATP Content in Rat Liver Mitochondria. J. Biochem. Mol. Toxicol. 2014, 28, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, J.E.; Jellum, E. The Identification and Metabolic Origin of 2-Furoylglycine and 2,5-Furandicarboxylic Acid in Human Urine. Clin. Chim. Acta 1972, 41, 199–207. [Google Scholar] [CrossRef]
- Mullur, R.; Liu, Y.Y.; Brent, G.A. Thyroid Hormone Regulation of Metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef] [PubMed]
- Pekary, A.E.; Sattin, A. Regulation of TRH and TRH-Related Peptides in Rat Brain by Thyroid and Steroid Hormones. Peptides 2001, 22, 1161–1173. [Google Scholar] [CrossRef]
- Hollenberg, A.N.; Monden, T.; Flynn, T.R.; Boers, M.E.; Cohen, O.; Wondisford, F.E. The Human Thyrotropin-Releasing Hormone Gene Is Regulated by Thyroid Hormone through Two Distinct Classes of Negative Thyroid Hormone Response Elements. Mol. Endocrinol. 1995, 9, 540–550. [Google Scholar] [CrossRef]
- Akinsanya, K.O.; Ghatei, M.A.; Bloom, S.R. Gonadal Steroids Regulate Rat Anterior Pituitary Levels of TSH-Releasing Hormone- and Pyroglutamyl-Glutamyl-Proline Amide-like Immunoreactivity. Endocrinology 1995, 136, 734–740. [Google Scholar] [CrossRef]
- Bennett, G.W.; Marsden, C.A.; Fone, K.C.F.; Johnson, J.V.; Heal, D.J. TRH-Catecholamine Interactions in Brain and Spinal Cord. Ann. N. Y. Acad. Sci. 1989, 553, 106–120. [Google Scholar] [CrossRef]
- Haus, E. Chronobiology in the Endocrine System. Adv. Drug Deliv. Rev. 2007, 59, 985–1014. [Google Scholar] [CrossRef]
- Breese, G.R.; Mueller, R.A.; Mailman, R.B.; Frye, G.D. Effects of TRH on Central Nervous System Function. Prog. Clin. Biol. Res. 1981, 68, 99–116. [Google Scholar]
- Mahmood, T.; Qureshi, I.Z.; Iqbal, M.J. Histopathological and biochemical changes in rat thyroid following acute exposure to hexavalent chromium. Histol. Histopathol. 2010, 25, 1355–1370. [Google Scholar] [PubMed]
- Li, Y.; Zhao, Y.; Deng, H.; Chen, A.; Chai, L. Endocrine Disruption, Oxidative Stress and Lipometabolic Disturbance of Bufo Gargarizans Embryos Exposed to Hexavalent Chromium. Ecotoxicol. Environ. Saf. 2018, 166, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, S.; Göethel, G.; Gauer, B.; Sauer, E.; Nardi, J.; Cestonaro, L.; Correia, D.; Peruzzi, C.; Mota, L.; Machry, R.V.; et al. Exposure to Environment Chemicals and Its Possible Role in Endocrine Disruption of Children from a Rural Area. Environ. Res. 2018, 167, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, S.; Choi, S.; Lee, I.; Moon, M.K.; Choi, K.; Park, Y.J.; Cho, Y.H.; Kwon, Y.M.; Yoo, J.; et al. Association of Exposure to Polycyclic Aromatic Hydrocarbons and Heavy Metals with Thyroid Hormones in General Adult Population and Potential Mechanisms. Sci. Total Environ. 2021, 762, 144227. [Google Scholar] [CrossRef] [PubMed]
- Motofei, I.G. A Dual Physiological Character for Cerebral Mechanisms of Sexuality and Cognition: Common Somatic Peripheral Afferents. BJU Int. 2011, 108, 1634–1639. [Google Scholar] [CrossRef]
- Henderson, B.E.; Ponder, B.A.J.; Ross, R.K. Hormones, Genes, and Cancer; Oxford University Press: Oxford, UK, 2003; pp. 1–484. [Google Scholar]
- Stárka, L. Epitestosterone. J. Steroid Biochem. Mol. Biol. 2003, 87, 27–34. [Google Scholar] [CrossRef]
- Aruldhas, M.M.; Subramanian, S.; Sekhar, P.; Hasan, G.C.; Govindarajulu, P.; Akbarsha, M.A. Microcanalization in the Epididymis to Overcome Ductal Obstruction Caused by Chronic Exposure to Chromium—A Study in the Mature Bonnet Monkey (Macaca radiata Geoffroy). Reproduction 2004, 128, 127–137. [Google Scholar] [CrossRef]
- Aruldhas, M.M.; Subramanian, S.; Sekar, P.; Vengatesh, G.; Chandrahasan, G.; Govindarajulu, P.; Akbarsha, M.A. Chronic Chromium Exposure-Induced Changes in Testicular Histoarchitecture Are Associated with Oxidative Stress: Study in a Non-Human Primate (Macaca radiata Geoffroy). Hum. Reprod. 2005, 20, 2801–2813. [Google Scholar] [CrossRef]
- Aruldhas, M.; Subramanian, S.; Sekhar, P.; Vengatesh, G.; Govindarajulu, P.; Akbarsha, M. In Vivo Spermatotoxic Effect of Chromium as Reflected in the Epididymal Epithelial Principal Cells, Basal Cells, and Intraepithelial Macrophages of a Nonhuman Primate (Macaca radiata Geoffroy). Fertil. Steril. 2006, 86, 1097–1105. [Google Scholar] [CrossRef]
- Subramanian, S.; Rajendiran, G.; Sekhar, P.; Gowri, C.; Govindarajulu, P.; Aruldhas, M.M. Reproductive Toxicity of Chromium in Adult Bonnet Monkeys (Macaca radiata Geoffrey). Reversible Oxidative Stress in the Semen. Toxicol. Appl. Pharmacol. 2006, 215, 237–249. [Google Scholar] [CrossRef]
- Danielsson, B.R.G.; Dencker, L.; Lindgren, A.; Tjälve, H. Accumulation of Toxic Metals in Male Reproduction Organs. In Disease, Metabolism and Reproduction in the Toxic Response to Drugs and Other Chemicals; Chambers, P.L., Preziosi, P., Chambers, C.M., Eds.; Archives of Toxicology; Springer Berlin Heidelberg: Berlin/Heidelberg, Germany, 1984; pp. 177–180. [Google Scholar] [CrossRef]
- Sutherland, J.E.; Zhitkovich, A.; Kluz, T.; Costa, M. Rats Retain Chromium in Tissues Following Chronic Ingestion of Drinking Water Containing Hexavalent Chromium. Biol. Trace Elem. Res. 2000, 74, 41–54. [Google Scholar] [CrossRef]
- Sekar, P.; Vengatesh, G.; Kumar, M.K.; Balaji, S.; Akbarsha, M.A.; Aruldhas, M.M. Impact of gestational and lactational exposure to hexavalent chromium on steroidogenic compartment of post-natal rat testis. J. Endocrinol. Reprod. 2011, 15, 15–26. [Google Scholar]
- Navin, A.K.; Aruldhas, M.M.; Navaneethabalakrishnan, S.; Mani, K.; Michael, F.M.; Srinivasan, N.; Banu, S.K. Prenatal Exposure to Hexavalent Chromium Disrupts Testicular Steroidogenic Pathway in Peripubertal F1 Rats. Reprod. Toxicol. 2021, 101, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Landry, A.P.; Ding, H. The Mitochondrial Outer Membrane Protein MitoNEET Is a Redox Enzyme Catalyzing Electron Transfer from FMNH2 to Oxygen or Ubiquinone. J. Biol. Chem. 2017, 29, 10061–10067. [Google Scholar] [CrossRef]
- Linnane, A.W.; Kios, M.; Vitetta, L. Coenzyme Q10—Its Role as a Prooxidant in the Formation of Superoxide Anion/Hydrogen Peroxide and the Regulation of the Metabolome. Mitochondrion 2007, 7, 51–61. [Google Scholar] [CrossRef]
- Littarru, G.P.; Tiano, L. Bioenergetic and Antioxidant Properties of Coenzyme Q10: Recent Developments. Mol. Biotechnol. 2007, 37, 31–37. [Google Scholar] [CrossRef]
- Crane, F.L. Biochemical Functions of Coenzyme Q10. J. Am. Coll. Nutr. 2001, 20, 591–598. [Google Scholar] [CrossRef]
- Hargreaves, I.P. Ubiquinone: Cholesterol’s Reclusive Cousin. Ann. Clin. Biochem. 2003, 40, 207–218. [Google Scholar] [CrossRef]
- Rodick, T.C.; Seibels, D.R.; Babu, J.R.; Huggins, K.W.; Ren, G.; Mathews, S.T. Potential Role of Coenzyme Q10 in Health and Disease Conditions. Nutr. Diet. Suppl. 2018, 10, 1–11. [Google Scholar] [CrossRef]
- Hargreaves, I.; Heaton, R.A.; Mantle, D. Disorders of Human Coenzyme Q10 Metabolism: An Overview. Int. J. Mol. Sci. 2020, 21, 6695. [Google Scholar] [CrossRef]
- Gutierrez-Mariscal, F.M.; Yubero-Serrano, E.M.; Villalba, J.M.; Lopez-Miranda, J. Coenzyme Q10: From Bench to Clinic in Aging Diseases, a Translational Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2240–2257. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, G.M.; El-Sayed, E.S.M.; Abo-Salem, O.M. Effect of Lead Toxicity on Coenzyme Q Levels in Rat Tissues. Food Chem. Toxicol. 2010, 48, 1753–1756. [Google Scholar] [CrossRef] [PubMed]
- Sijko, M.; Janasik, B.; Wąsowicz, W.; Kozłowska, L. Can the Effects of Chromium Compounds Exposure be Modulated by Vitamins and Microelements? Int. J. Occup. Med. Environ. Health 2021, 34, 461–490. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.S. Protective Effect of Vitamin B6 in Chromium-Induced Oxidative Stress in Liver. J. Appl. Toxicol. 2005, 25, 440–443. [Google Scholar] [CrossRef]
- Yang, T.H.; Hu, M.L. Intracellular Levels of S-Adenosylhomocysteine but Not Homocysteine Are Highly Correlated to the Expression of Nm23-H1 and the Level of 5-Methyldeoxycytidine in Human Hepatoma Cells with Different Invasion Activities. Nutr. Cancer 2006, 55, 224–231. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, S.; Xie, Z.; Blum, W.; Perrotti, D.; Paschka, P.; Klisovic, R.; Byrd, J.; Chan, K.K.; Marcucci, G. Characterization of in Vitro and in Vivo Hypomethylating Effects of Decitabine in Acute Myeloid Leukemia by a Rapid, Specific and Sensitive LC-MS/MS Method. Nucleic Acids Res. 2007, 35, 31. [Google Scholar] [CrossRef]
- Cho, S.H.; Jung, B.H.; Lee, S.H.; Lee, W.Y.; Kong, G.; Chung, B.C. Direct Determination of Nucleosides in the Urine of Patients with Breast Cancer Using Column-Switching Liquid Chromatography-Tandem Mass Spectrometry. Biomed. Chromatogr. 2006, 20, 1229–1236. [Google Scholar] [CrossRef]
- Kanduc, D. 1,7-Dimethylguanosine Formation in TRNA Enzymatic Methylation. Boll. Soc. Ital. Biol. Sper. 1982, 58, 453–456. [Google Scholar]
- Niwa, T.; Takeda, N.; Yoshizumi, H. RNA Metabolism in Uremic Patients: Accumulation of Modified Ribonucleosides in Uremic Serum. Technical Note. Kidney Int. 1998, 53, 1801–1806. [Google Scholar] [CrossRef]
- Mathé, E.A.; Patterson, A.D.; Haznadar, M.; Manna, S.K.; Krausz, K.W.; Bowman, E.D.; Shields, P.G.; Idle, J.R.; Smith, P.B.; Anami, K.; et al. Noninvasive Urinary Metabolomic Profiling Identifies Diagnostic and Prognostic Markers in Lung Cancer. Cancer Res. 2014, 74, 3259–3270. [Google Scholar] [CrossRef]
- Kable, E.P.W.; Parsons, P.G. Melanin Synthesis and the Action of L-Dopa and 3,4-Dihydroxybenzylamine in Human Melanoma Cells. Cancer Chemother. Pharmacol. 1989, 23, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Koshimura, I.; Imai, H.; Hidano, T.; Endo, K.; Mochizuki, H.; Kondo, T.; Mizuno, Y. Dimethoxyphenylethylamine and Tetrahydropapaverine Are Toxic to the Nigrostriatal System. Brain Res. 1997, 773, 108–116. [Google Scholar] [CrossRef]
- Goto, K.; Mochizuki, H.; Hattori, T.; Nakamura, N.; Mizuno, Y. Neurotoxic Effects of Papaverine, Tetrahydropapaverine and Dimethoxyphenylethylamine on Dopaminergic Neurons in Ventral Mesencephalic-Striatal Co-Culture. Brain Res. 1997, 754, 260–268. [Google Scholar] [CrossRef]
- Lümmen, P. Complex I Inhibitors as Insecticides and Acaricides. Biochim. Biophys. Acta 1998, 1364, 287–296. [Google Scholar] [CrossRef]
- Greenamyre, J.T.; Sherer, T.B.; Betarbet, R.; Panov, A.V. Complex I and Parkinson’s Disease. IUBMB Life 2001, 52, 135–141. [Google Scholar] [CrossRef]
- Dodd, D.; Spitzer, M.H.; Van Treuren, W.; Merrill, B.D.; Hryckowian, A.J.; Higginbottom, S.K.; Le, A.; Cowan, T.M.; Nolan, G.P.; Fischbach, M.A.; et al. A Gut Bacterial Pathway Metabolizes Aromatic Amino Acids into Nine Circulating Metabolites. Nature 2017, 551, 648–652. [Google Scholar] [CrossRef]
- Jellet, J.J.; Forrest, T.P.; Macdonald, I.A.; Marrie, T.J.; Holdeman, L.V. Production of Indole-3-Propanoic Acid and 3-(p-Hydroxyphenyl)Propanoic Acid by Clostridium Sporogenes: A Convenient Thin-Layer Chromatography Detection System. Can. J. Microbiol. 1980, 26, 448–453. [Google Scholar] [CrossRef]
- Karbownik, M.; Reiter, R.J.; Garcia, J.J.; Cabrera, J.; Burkhardt, S.; Osuna, C.; Lewinski, A. Indole-3-Propionic Acid, a Melatonin-Related Molecule, Protects Hepatic Microsomal Membranes from Iron-Induced Oxidative Damage: Relevance to Cancer Reduction. J. Cell. Biochem. 2001, 81, 507–513. [Google Scholar] [CrossRef]
- Gaetani, L.; Boscaro, F.; Pieraccini, G.; Calabresi, P.; Romani, L.; Di Filippo, M.; Zelante, T. Host and Microbial Tryptophan Metabolic Profiling in Multiple Sclerosis. Front. Immunol. 2020, 11, 157. [Google Scholar] [CrossRef]
- Diez-Fernandez, C.; Hertig, D.; Loup, M.; Diserens, G.; Henry, H.; Vermathen, P.; Nuoffer, J.; Häberle, J.; Braissant, O. Argininosuccinate Neurotoxicity and Prevention by Creatine in Argininosuccinate Lyase Deficiency: An in Vitro Study in Rat Three-dimensional Organotypic Brain Cell Cultures. J. Inherit. Metab. Dis. 2019, 42, 1077–1087. [Google Scholar] [CrossRef]
- Baruteau, J.; Perocheau, D.P.; Hanley, J.; Lorvellec, M.; Rocha-Ferreira, E.; Karda, R.; Ng, J.; Suff, N.; Diaz, J.A.; Rahim, A.A.; et al. Argininosuccinic Aciduria Fosters Neuronal Nitrosative Stress Reversed by Asl Gene Transfer. Nat. Commun. 2018, 9, 3505. [Google Scholar] [CrossRef] [PubMed]
- Nagamani, S.C.S.; Erez, A.; Lee, B. Argininosuccinate Lyase Deficiency. Genet. Med. 2012, 14, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Seminotti, B.; da Silva, J.C.; Ribeiro, R.T.; Leipnitz, G.; Wajner, M. Free Radical Scavengers Prevent Argininosuccinic Acid-Induced Oxidative Stress in the Brain of Developing Rats: A New Adjuvant Therapy for Argininosuccinate Lyase Deficiency? Mol. Neurobiol. 2020, 57, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.H.; Wang, K.C.; Tian, T.F.; Tsai, M.H.; Chiung, Y.M.; Hsiech, C.M.; Tsai, S.J.; Wang, S.Y.; Tsai, D.M.; Huang, C.C.; et al. Metabolomic Characterization of Laborers Exposed to Welding Fumes. Chem. Res. Toxicol. 2012, 25, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Santonen, T.; Alimonti, A.; Bocca, B.; Duca, R.C.; Galea, K.S.; Godderis, L.; Göen, T.; Gomes, B.; Hanser, O.; Iavicoli, I.; et al. Setting up a Collaborative European Human Biological Monitoring Study on Occupational Exposure to Hexavalent Chromium. Environ. Res. 2019, 177, 108583. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Biological Monitoring of Chemical Exposure in the Workplace; WHO: Geneva, Switzerland, 1996; pp. 1–314. [Google Scholar]
- Esteban López, M.; Göen, T.; Mol, H.; Nübler, S.; Haji-Abbas-Zarrabi, K.; Koch, H.M.; Kasper-Sonnenberg, M.; Dvorakova, D.; Hajslova, J.; Antignac, J.P.; et al. The European Human Biomonitoring Platform—Design and Implementation of a Laboratory Quality Assurance/Quality Control (QA/QC) Programme for Selected Priority Chemicals. Int. J. Hyg. Environ. Health 2021, 234, 113740. [Google Scholar] [CrossRef]
- Southam, A.D.; Haglington, L.D.; Najdekr, L.; Jankevics, A.; Weber, R.J.M.; Dunn, W.B. Assessment of Human Plasma and Urine Sample Preparation for Reproducible and High-Throughput UHPLC-MS Clinical Metabolic Phenotyping. Analyst 2020, 145, 6511–6523. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The Human Metabolome Database for 2018. Nucleic Acids Res. 2018, 46, 608–617. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Wang, S.; Chen, X.; Du, D.; Zheng, W.; Hu, L.; Yang, H.; Cheng, J.; Gong, M. MetaboGroup S: A Group Entropy-Based Web Platform for Evaluating Normalization Methods in Blood Metabolomics Data from Maintenance Hemodialysis Patients. Anal. Chem. 2018, 90, 11124–11130. [Google Scholar] [CrossRef]
- Hur, M.; Campbell, A.A.; Almeida-de-Macedo, M.; Li, L.; Ransom, N.; Jose, A.; Crispin, M.; Nikolau, B.J.; Wurtele, E.S. A Global Approach to Analysis and Interpretation of Metabolic Data for Plant Natural Product Discovery. Nat. Prod. Rep. 2013, 30, 565. [Google Scholar] [CrossRef] [PubMed]
- Zuba, D.; Parczewski, A. Chemometria w Analityce; Instytut Ekspertyz Sadowych: Krakow, Poland, 2008; pp. 1–328. [Google Scholar]
- Jolliffe, I.T. Principal Component Analysis, 2nd ed.; Springer: New York, NY, USA, 2002; pp. 1–518. [Google Scholar]
- Praus, P. SVD-based principal component analysis of geochemical data. Cent. Eur. J. Chem. 2005, 3, 731–741. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Principal component analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Mazerski, J. Podstawy Chemometrii; Wydawnictwo Politechniki Gdańskiej: Gdansk, Poland, 2000; pp. 1–189. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country/ Groups | All Workers (n = 220) | WW (n = 101) | WCP (n = 61) | WST (n = 58) | All Controls (n = 102) | WCC (n = 33) | OCC (n = 69) |
|---|---|---|---|---|---|---|---|
| Poland | 50 | 49 | - | 1 | 45 | 13 | 32 |
| Portugal | 45 | 3 | 7 | 35 | 24 | 2 | 22 |
| Finland | 46 | 23 | 18 | 5 | 21 | 6 | 15 |
| Belgium | 59 | 26 | 16 | 17 | 7 | 7 | - |
| The Netherlands | 20 | - | 20 | - | 5 | 5 | - |
| Parameter | All Workers (n = 220) | WW (n = 101) | WCP (n = 61) | WST (n = 58) | All Controls (n = 102) | WCC (n = 33) | OCC (n = 69) |
|---|---|---|---|---|---|---|---|
| Age | 41.7 ± 11.0 | 39.1 ± 10.7 | 43.8 ± 12.2 | 43.9 ± 9.4 | 44.8 ± 10.1 | 44.0 ± 9.0 | 45.1 ± 11.0 |
| Height (cm) | 178.0 ± 6.4 | 177.7 ± 6.2 | 179.7 ± 6.5 | 176.6 ± 6.2 | 178.4 ± 6.9 | 180.7 ± 5.7 | 177.3 ± 7.1 |
| Body mass (kg) | 85.6 ± 13.6 | 85.1 ± 13.8 | 88.4 ± 15.1 | 83.6 ± 10.8 | 85.8 ± 13.0 | 91.5 ± 12.4 | 83.0 ± 12.4 |
| BMI (kg/m2) | 27.0 ± 4.0 | 26.9 ± 3.8 | 27.4 ± 4.8 | 26.8 ± 3.4 | 27.0 ± 3.8 | 28.0 ± 3.8 | 26.5 ± 3.7 |
| Smoking status | |||||||
| No (%) | 52.1 | 59.4 | 37.7 | 54.5 | 75.2 | 81.3 | 72.5 |
| Yes (%) | 33.6 | 25.7 | 42.6 | 38.2 | 15.8 | 9.4 | 18.8 |
| Former smoker (%) | 14.3 | 14.9 | 19.7 | 7.3 | 8.9 | 9.4 | 8.7 |
| Alcohol use | |||||||
| No (%) | 15.7 | 10.9 | 9.8 | 30.9 | 26.0 | 6.3 | 35.3 |
| Yes (%) | 84.3 | 89.0 | 90.0 | 69.0 | 74.0 | 93.7 | 64.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozłowska, L.; Santonen, T.; Duca, R.C.; Godderis, L.; Jagiello, K.; Janasik, B.; Van Nieuwenhuyse, A.; Poels, K.; Puzyn, T.; Scheepers, P.T.J.; et al. HBM4EU Chromates Study: Urinary Metabolomics Study of Workers Exposed to Hexavalent Chromium. Metabolites 2022, 12, 362. https://doi.org/10.3390/metabo12040362
Kozłowska L, Santonen T, Duca RC, Godderis L, Jagiello K, Janasik B, Van Nieuwenhuyse A, Poels K, Puzyn T, Scheepers PTJ, et al. HBM4EU Chromates Study: Urinary Metabolomics Study of Workers Exposed to Hexavalent Chromium. Metabolites. 2022; 12(4):362. https://doi.org/10.3390/metabo12040362
Chicago/Turabian StyleKozłowska, Lucyna, Tiina Santonen, Radu Corneliu Duca, Lode Godderis, Karolina Jagiello, Beata Janasik, An Van Nieuwenhuyse, Katrien Poels, Tomasz Puzyn, Paul T. J. Scheepers, and et al. 2022. "HBM4EU Chromates Study: Urinary Metabolomics Study of Workers Exposed to Hexavalent Chromium" Metabolites 12, no. 4: 362. https://doi.org/10.3390/metabo12040362
APA StyleKozłowska, L., Santonen, T., Duca, R. C., Godderis, L., Jagiello, K., Janasik, B., Van Nieuwenhuyse, A., Poels, K., Puzyn, T., Scheepers, P. T. J., Sijko, M., Silva, M. J., Sosnowska, A., Viegas, S., Verdonck, J., Wąsowicz, W., on behalf of HBM4EU Chromates Study Team, & on behalf of Statistical Team. (2022). HBM4EU Chromates Study: Urinary Metabolomics Study of Workers Exposed to Hexavalent Chromium. Metabolites, 12(4), 362. https://doi.org/10.3390/metabo12040362